GUCY2F antibody
Cat. No. GTX14789
Cat. No. GTX14789
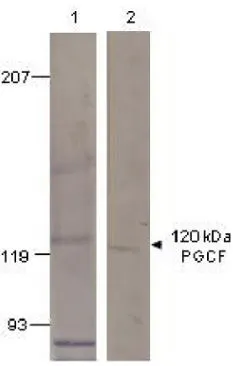
GTX14789 WB Image
WB analysis of PGCF recombinant protein using GTX14789 GUCY2F antibody.
Lane 1 : Pre-immune serum
Lane 2 : Primary antibody
Dilution : 1:500
1 / 2
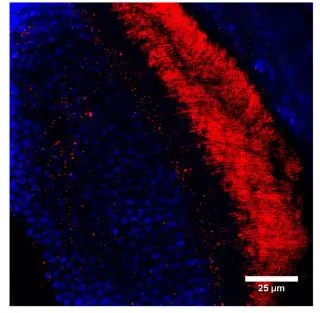
GTX14789 IHC Image
IHC analysis of mouse retina tissue (photoreceptor outer segments) using GTX14789 GUCY2F antibody.
Red : Primary antibody
Blue : Nuclei
2 / 2
-
HostRabbit
-
ClonalityPolyclonal
-
IsotypeIgG
-
ApplicationsWB ICC/IF ELISA IHC
-
ReactivityHuman, Mouse, Rat